
Emilie Fairfax , Chris Jordan
Castor : le plan d’action climat américain en eau douce
Traduit par MAPCa
AVANT PROPOS
Cet article d’Emily Fairfax et Chris Jordan est un texte fondateur pour le mouvement d’alliance avec le peuple castor. Publié en 2022 par ces deux chercheurs éminents en écohydrologie et en biologie, dans une revue scientifique peer-reviewed, il a donné sa plateforme théorique et scientifique au mouvement de la beaver based restoration, la restauration fondée sur les castors (abrégée ici parfois en « restauration castor »), qui existait sous plusieurs formes et manifestations aux États-Unis. Fairfax et Jordan incluent dans cette approche trois types d’action :
- la favorisation de la présence des castors dans les milieux rivière pour qu’ils produisent leurs effets écologiques (contre la tendance traditionnelle de les piéger pour les mettre à mort, dominantes aux Etats Unis où l’animal n’est pas protégé par la loi) ;
- la relocalisation de castors (déplacer des individus qui posaient problèmes dans des milieux où ils peuvent jouer leur rôle, et réintroduire des castors là où le milieu a besoin d’eux) ;
- la régénération low tech fondée sur les processus, telle qu’elle est théorisée par Joe Wheaton et ses collègues à l’Université d’Utah.
Cet article est précieux car sa bibliographie scientifique est une mine d’informations qui répond à presque toutes les questions que l’on peut se poser sur les rapports entre castor, milieu, et changement climatique, et qui donne des arguments et des faits quantifiés pour soutenir la pertinence et l’importance d’une alliance avec le peuple castor.
Dans le texte Fairfax et Jordan mobilisent spontanément les concepts de « service écosystémique », et de « restauration » : ce sont des notions que nous essayons de critiquer et dépasser dans le MAPCa, comme c’est formulé dans les argumentations de Rendre l’eau à la terre.
Ces nuances terminologiques sont importantes dans certains contextes, mais elles ne sont pas décisives ici, et n’entament en rien l’importance de ce texte pour penser un autre rapport aux rivières et aux hydrosystèmes, en changeant nos relations avec la vie non humaines qui les anime.
Néanmoins c’est un texte technique, impliquant une grande culture en écohydrologie et hydromorphologie, et il peut être utile pour le décrypter de s’appuyer sur une introduction philosophique des enjeux, parfois obscurcis par le langage technique des sciences de l’eau. Vous pouvez lire cette introduction avant l’article, après, ou jamais, elle n’est pas nécessaire pour comprendre le texte mais elle tente de révéler ces enjeux écologiques profonds.
Introduction aux enjeux écologiques et philosophiques du « plan d’action castor pour le climat en eau douce »
La puissance politique et écologique de cet article scientifique d’Emily Fairfax et Chris Jordan, est qu’il rend visible un pan invisible et majeur de notre monde commun, de nos milieux vivants. Ce pan invisible, ce haut lieu où la vie prospère et rayonne, qui est partout autour de nous, mais que nous ne voyons pas, est formulé dans ce texte par la formule obscure et peu spectaculaire de « connected floodplains ». Dans la question des relations entre l’eau et la vie sur les continents, quelque chose d’essentiel se joue là, pour faire face au changement climatique, et amplifier la vie, et nous ne le voyons pas, comme société ni comme individus.
C’est un concept hydromorphologique d’allure abstraite, peu évocateur, mais qui, lorsqu’on comprend pleinement ses implications, transforme la conception des milieux dans lesquels nous vivons.
Une manière de la raconter, c’est de restituer l’histoire par laquelle la vie complexe est sortie des eaux il y a quelques 400 millions d’années pour s’établir sur la terre ferme, sur les continents. Elle n’a pu se déployer et prospérer qu’en multipliant les dispositifs, les inventions évolutionnaires et écologiques pour garder l’eau avec elle : pour labyrinthiser l’eau en elle et autour d’elle.
Imaginez que les continents, avant la sortie des eaux, étaient comme un désert. La vie a contribué, en ralentissant l’eau, en la ramifiant dans les écosystèmes, à en faire des oasis.
Avance rapide jusqu’au 19ème siècle : l’ère moderne se définit comme l’ère du drainage. L’ère du drainage a consisté à accélérer et optimiser l’évacuation de l’eau en dehors des milieux, à des fins agricoles et urbaines. Si on comprend bien l’histoire du temps profond, il est clair que ce faisant, on s’est opposé aux inventions du vivant de terre ferme qui avaient pour vocation de faire prospérer la vie en gardant l’eau avec elle, car la vie a besoin de l’eau.
Qu’est-ce que c’est qu’une « connected floodplain » sous cet angle ? C’est l’idée qu’en vérité, un cours d’eau n’est pas le trait bleu sur la carte. Il est vitalement connecté à articulé à l’ensemble des milieux du fond de vallée, et même du bassin versant, dans lesquels la vie s’active et module le comportement de l’eau qui vient de la pluie et la neige. Il est vitalement articulé avec sa nappe d’accompagnement, en surface, qui elle-même peut alimenter des nappes plus profondes.
Ce que nous avons fait, c’est massivement déconnecter la plaine alluviale, c’est-à-dire le fond de vallée, qui a auparavant impliqué des forêts, des prairies humides, des marais, des zones humides, des nappes – avec les cours d’eau eux-mêmes. Cette déconnexion se manifeste par l’incision, la simplification des rivières à chenaux multiples en chenaux uniques, l’homogénéisation des habitats, et la déconnexion entre le cours d’eau et la nappe, qui n’est plus autant capable d’activer ses échanges nappe-rivière. Cette déconnexion tend à aggraver les effets du changement climatique par des sécheresses plus longues et intenses, des inondations plus fréquentes, des feux plus intenses et une accélération de l’érosion de la biodiversité.
Imaginez qu’auparavant, le sol du fond de vallée et du bassin versant était une gigantesque éponge qui gardait l’eau tranquillement, alimentant la nappe qui soutient les basses eaux de la rivière, augmentant le temps de résidence de l’eau dans les milieux, la faisait tourner dans les système racinaire, dans les champignons, dans la faune des sols, la ralentissait dans les rivières, les marécages, les prairies humides, la gardait dans les embâcles, les ouvrages de castor. Nous avons massivement fragilisé cette capacité des milieux à garder l’eau avec eux, en favorisant systématiquement les forces de ruissellement au détriment des forces d’infiltration : en drainant les zones humides, en bétonnant via l’étalement urbain, en vidant la matière organique des sols, en enlevant le couvert végétal dans les milieux agricoles, en enlevant la matière organique qui se gorge d’eau dans les sols. Et ce faisant, nous avons massivement déconnecté l’eau et la terre : les écoulements de l’eau qui, avec la pluie, prennent 1 000 chemins dans le chevelu pour se retrouver dans les rivières – et les milieux vivants : des zones hyporhéiques où se trouvent les frayères de poissons migrateurs, des ripisylves, des zones humides aux bras morts de la rivière où vivent des milliers d’espèces terrestres et aquatiques, champignons, animaux et végétaux.
C’est comme si nous avions déconnecté le système veineux et le corps. Le corps, c’est l’ensemble des milieux vivants de fond de vallée. Le système veineux, c’est la part d’écoulement rapide de l’hydrosystème, incluant le chevelu par lequel les petits écoulements, à chaque pluie, se rejoignent en ramification pour atteindre les rivières, mais aussi tous les visages lents de l’eau bleue, dans les lacs. Un des aspects clé de l’hydrosystème malade aujourd’hui est que le cours d’eau est déconnecté des nappes. Nous avons coupé toutes les connexions entre les mille tissages de l’eau dans les milieux vivants : celui entre la nappe et la rivière est parmi les plus graves et les plus important à reconstituer chaque fois que c’est possible. C’est comme si nous avions débranché systématiquement le système sanguin du corps lui-même et que le corps restait sec, non irrigué, non hydraté, non nourri, parce qu’il est déconnecté du réseau qui fait circuler l’eau.
Nous avons pris l’eau à la terre, nous avons déconnecté la terre et l’eau.
Répondre à ce problème, c’est ce que veut dire rendre l’eau à la terre.
C’est ce que veut dire rendre l’eau à la vie : c’est-à-dire reconstituer toutes les connexions par lesquelles l’eau dans les bassins versants, dans les rivières, est connectée au milieu vivant et hydrate la vie, fait fonctionner la vie, fait prospérer l
La grande découverte que développent ici Emily Fairfax et Chris Jordan, c’est qu’il y a un animal dont l’activité spontanée depuis des millions d’années reconnecte la terre et l’eau, reconnecte les plaines alluviales à leurs cours d’eau, les cours d’eau à leur nappes, l’eau et la vie, les flux et les éponges. Ils ont pu scientifiquement documenter de manière minutieuse et globale l’ensemble des effets extraordinaires de l’action de ces animaux : d’atténuation des crues, de soutien d’étiage, d’atténuation des effets des sécheresses, de soutien à la diversité des espèces, de tampons à l’égard des mégafeux, d’adaptation au changement climatique, de robustesse et de résilience des écosystèmes.
Tout ça à partir d’un principe simple, un principe d’action dans le rapport à l’eau. Nous avons déconnecté systématiquement l’hydrosystème et les milieux vivants. L’enjeu est de reconnecter à chaque fois que possible les flux de l’eau et les milieux vivants. Reconnecter à chaque fois que c’est possible à l’intérieur de l’hydrosystème toutes les connexions qui ont été coupées : entre rivière, sol et nappe, rivière et plaine alluviale, rivière et zone humide…
L’hydrosystème, c’est précisément ce système qui englobe les cours d’eau, des petits chevelus aux grands fleuves, des têtes de bassin versant à la mer (l’eau bleue) d’un côté et de l’autre, les milieux vivants, les prairies, les forêts de rive, les zones humides (l’eau verte). Dans l’hydrosystème, nous avons coupé de manière systématique les connexions entre l’eau et les milieux vivants. L’enjeu est de régénérer les connexions entre la terre et l’eau, entre le corps de la terre que sont les milieux vivants et les veines d’eau, à chaque fois que c’est possible dans l’hydrosystème.
C’est ça que le castor permet de penser et c’est vers là qu’il permet d’imaginer des formes émancipatrices d’action. Ce n’est pas vouer un culte au castor, ou l’instrumentaliser pour résoudre nos problèmes. Ce qui a lieu dans la pensée de Fairfax et Jordan, c’est une intercession fascinante : comment leur enquête scientifique sur les effets de l’action du castor a changé leur conception du rapport des humains à l’eau, et le sens de notre action possible. C‘est la médiation par la compréhension des effets de l’action d’un autre animal sur les milieux (reconnecter la terre vivante et l’eau), qui peut nous inspirer quant à comment régénérer nos milieux face aux bouleversements du changement climatique.
L’article mobilise beaucoup le concept de service écosystémique. C’est un concept technocratique qui, dans certaines arènes, est capable de jouer un rôle politique. Nous ne revendiquons pas ce concept parce que dans beaucoup de cas, il produit plus d’effets délétères que d’effets émancipateurs, notamment par ses effets de dépolitisation de la question des usages humains de la terre. On préfère parler d’alliance avec des non humains et on vous renvoie au livre Rendre l’eau à la terre (cf. pages 290 et 291) pour comprendre pourquoi.
De la même manière, l’article mobilise beaucoup le concept de « solution fondé sur la nature ». Ce concept est utile et défendable dans plusieurs contextes et arènes, notamment institutionnelles. Nous ne mobilisons pas non plus ce concept parce que d’un point de vue philosophique, il appartient souvent à un imaginaire ingénierique qui consiste à ne pas questionner la formulation même des problèmes et à penser le monde naturel sous un angle instrumental comme des solutions à des problèmes strictement humains. On préfère parler d’approche systémique de soin des milieux au service de la vie, et donc des sociétés humaines soutenables plutôt que de « solutions fondées sur la nature ».
La puissance de cet article, c’est de fonder scientifiquement sur une bibliographie extensive, précise et rigoureuse, l’esquisse d’un changement de paradigme dans le rapport à l’eau des rivières, des hydrosystèmes, dans lequel il s’agit de rendre visible quelque chose que nous n’avions pas vu : reconnecter les cours d’eau aux milieux vivants et à leur corps tout entier, les bassins versants. La veille, on n’était pas au courant que les milieux continentaux souffraient d’une déconnexion fondamentale dans l’hydrosystème entre les milieux vivants et les cours d’eau. Avec cet article, émerge simultanément un observable qui a l’air assez abstrait, mais qui est en fait très clair quand on le regarde précisément. Nous avons déconnecté les veines des continents et le corps de terre. Il est temps de rendre l’eau à la terre, et de rendre la terre à l’eau, au service de la vie pour restituer aux milieux vivants la robustesse dont ils ont besoin pour traverser le changement climatique de manière moins fragile et continuer à abriter la vie plurispécifique, à nous hydrater et nous nourrir nous aussi, qui en faisons partie, et à rendre habitables les milieux pour des sociétés humaines qui auraient enfin changé de cap.
Castor : le plan d’action climat américain en eau douce
Chris E. Jordan, Emily Fairfax Publié le 28 avril 2022
Résumé
Les rivières et les ruisseaux, lorsqu’ils sont entièrement connectés à leurs plaines alluviales, sont des systèmes naturellement résilients qui font de plus en plus partie du débat sur les solutions climatiques fondées sur la nature. La reconnexion des cours d’eau à leurs lits majeurs améliore la qualité et la quantité de l’eau, favorise la biodiversité et la conservation des espèces sensibles, augmente la résilience aux inondations, aux sécheresses et aux incendies, et renforce la séquestration du carbone. Cependant, si l’importance de la restauration des cours d’eau est évidente, la “restauration castor” (beaver-based restauration) – impliquant par exemple la coexistence avec les castors, leur relocalisation et le mimétisme de leurs effets écologiques – reste sous-utilisée malgré les nombreuses données démontrant son efficacité. Les perturbations liées au climat poussent activement les cours d’eau vers des états de plus en plus dégradés, et la fenêtre d’opportunité pour leur restauration ne restera pas ouverte éternellement. Par conséquent, c’est le moment idéal pour appliquer la science de la restauration fondée sur les processus, low tech, bas carbone et inspirée des castors pour grandir en résilience climatique à l’échelle du territoire. Tous les cours d’eau ne seront pas de bons candidats pour la “restauration castor” ou des processus naturels, mais nous avons les outils pour savoir lesquels le sont. Utilisons-les.
1 – Introduction : Les castors, le plan d’action climatique
La restauration low-tech fondée sur les processus – une série de pratiques simples et peu coûteuses axées sur la reconnexion des cours d’eau aux plaines alluviales – gagne rapidement du terrain face aux crises imminentes du climat et de la biodiversité (Ciotti et al, 2021 ; Davee et al., 2019 ; Davis et al., 2021 ; Johnson et al., 2019 ; Keeble-Toll, 2018 ; Munir & Westbrook, 2020 ; Pearce et al., 2021a, 2021b ; Silverman et al., 2019 ; Wade et al., 2020 ; Weber et al., 2017 ; Wheaton et al., 2019). Bien que la mise en œuvre de ces méthodes repose sur des fondements théoriques et techniques solides, le scepticisme persiste – en particulier en ce qui concerne l’efficacité des structures construites à la main, inspirées par les castors, et la coexistence avec les castors. En particulier, des publications récentes ont remis en question la possibilité d’obtenir des résultats à l’échelle du bassin versant par le biais des modifications créées par les castors ou d’ouvrages inspirés des castors (Nash et al., 2018 ; Nash et al., 2021 ; Pilliod et al., 2017). Ceci en dépit d’innombrables années de connaissances indigènes sur la gestion durable des rives et des castors (Albert & Trimble, 2000 ; Blackfeet Nation, 2018 ; Blackfeet Nation & Levitus, 2019 ; Feit, 1986 ; Gadgil et al, 1993 ; Keeble-Toll, 2018 ; Kimmerer, 2000 ; Kimmerer & Lake, 2001 ; Sherriff, 2021) et plus d’un siècle de données, d’expériences et d’analyses publiées (Ives, 1942 ; Morgan, 1868 ; Neff, 1957 ; Ruedemann & Schoonmaker, 1938 ; Seton, 1929) documentant une activité hyporhéique accrue (Briggs et al, 2013 ; Janzen & Westbrook, 2011 ; X. Wang et al., 2018), l’amélioration de la qualité de l’eau (Cornell et al., 2011 ; Lazar et al., 2015 ; Puttock et al., 2017, 2018 ; Shepherd & Nairn, 2020, 2021), la naturalisation de la dynamique des écoulements (Burchsted et al, 2010), l’échec des approches d’ingénierie traditionnelles de la restauration (D. M. Thompson & Stull, 2002), et la résilience aux incendies de forêt (Fairfax & Whittle, 2020 ; Foster et al., 2020 ; Weirich, 2021 ; Whipple, 2019). Les paysages fluviaux entièrement connectés aux plaines alluviales et occupés par les castors (Brazier et al., 2021 ; Larsen et al., 2021) sont des espaces de processus naturels que nous ne pouvons plus nous permettre d’ignorer.
Il peut sembler surprenant de dire que les castors sont un élément clé d’un plan national d’action climatique, mais la réalité est qu’ils représentent une force de 15 à 40 millions (Naiman et al., 1988) d’ingénieurs environnementaux hautement qualifiés. Nous ne pouvons plus nous permettre de travailler contre eux ; nous devons travailler avec eux. Dans la plupart des cas, la première étape consistera à amorcer le processus de restauration physique avant que les castors ne s’installent dans un hydrosystème, ce qui permettra aux processus de plaine alluviale de fonctionner (flux, espace, structure ; Beechie et al., 2010, Cluer & Thorne, 2014, Wheaton et al., 2019). Une intervention humaine peut être nécessaire pour restaurer les processus de plaine alluviale sévèrement dégradés jusqu’au stade où les castors et leur mimétisme peuvent être appliqués (par exemple, des chenaux profondément incisés, des perturbations continues des terres). Dans d’autres situations, la première étape peut consister en un changement de politique : par exemple, si les plaines alluviales sont intactes, mais que les méthodes de gestion des castors (par exemple, l’élimination des castors qui ont un impact sur le bâti) empêchent la présence d’une population suffisante pour permettre le rétablissement de ces paysages. Quel que soit notre rôle dans la conversation, la régénération des processus naturels ou inspirée des castors devrait être une stratégie principale pour retrouver des paysages fluviaux en bonne santé (Macfarlane et al., 2015 ; Pollock et al., 2015). Un cours d’eau où les castors prospèrent est un cours d’eau résilient et productif (Pollock et al., 2014). Des populations de castors florissantes peuvent être nos partenaires dans la lutte contre le changement climatique et un indicateur de notre progrès.

Restauration par la forme vs régénération en activant les processus naturels sur des zones plus ou moins contraintes. Aquarelle de Suzanne Husky inspirée d’un dessin de Damion Ciotti et Michael Pollock.
2 – La restauration des paysages fluviaux est une solution à portée de main
Le changement climatique amplifie les impacts de la dégradation des paysages fluviaux : des précipitations extrêmes plus fréquentes dans des lits de cours d’eau recalibrés entraînent davantage d’inondations (Stott, 2016) ; l’augmentation de la température de l’air et les conditions plus sèches stressent la végétation de fond de vallée déjà isolée des aquifères hyporhéiques, transformant les feux de forêt en ” méga-feux ” (Finco et al, 2012 ; Goss et al., 2020 ; Mori & Johnson, 2013 ; Swain, 2021 ; J. Williams, 2013 ; A. P. Williams et al., 2019) ; et les régimes hydrologiques centrés sur la neige qui se transforment en régimes hydrologiques pluviaux entraînent des débits de base plus faibles et plus chauds et une dégradation supplémentaire des conditions biotiques (Beechie et al., 2013). Cependant, nous ne développons pas de stratégies d’action climatique fondées sur la nature à l’échelle du paysage fluvial (Skidmore & Wheaton, 2022). La restauration de la connectivité et de la fonctionnalité des plaines alluviales est à la fois une stratégie d’atténuation et d’adaptation au changement climatique, car elle inverse la dégradation et restaure la résilience naturelle (Johnson et al., 2019 ; Pollock et al., 2015 ; Silverman et al., 2019 ; Wheaton et al., 2019). La résilience naturelle du paysage fluvial n’est possible que par une restauration des processus de la plaine alluviale (Cluer & Thorne, 2014), et non par l’ingénierie ou l’imposition d’une forme (ELI, 2016).
Selon la dernière évaluation de l’EPA, 25 à 50 % des eaux vives des États-Unis (CONUS uniquement) sont en mauvais état ( USEPA, 2013). L’activité humaine a considérablement réduit la connectivité des plaines alluviales à travers le continent, en convertissant les zones humides s’étendant sur toute la vallée en corridors rivulaires étroits. Par exemple, dans la vallée de Sacramento en Californie, les forêts riveraines des plaines alluviales bien connectées ont été réduites de 24 % à moins de 0,5 % de la surface des terres (Sands & Howe, 1977). Nos activités au cours des deux derniers siècles ont réduit la superficie des plaines alluviales actives d’un ordre de grandeur et dégradé la moitié des eaux courantes des États-Unis. La dégradation humaine des paysages fluviaux, si elle n’est pas maîtrisée, crée des cycles de rétroaction positive de dégradation supplémentaire dans un climat changeant (Figure 1). Cependant, la restauration fondée sur le castor offre de grandes possibilités pour activer des cycles d’auto aggradation positifs et améliorer la résilience des paysages fluviaux.
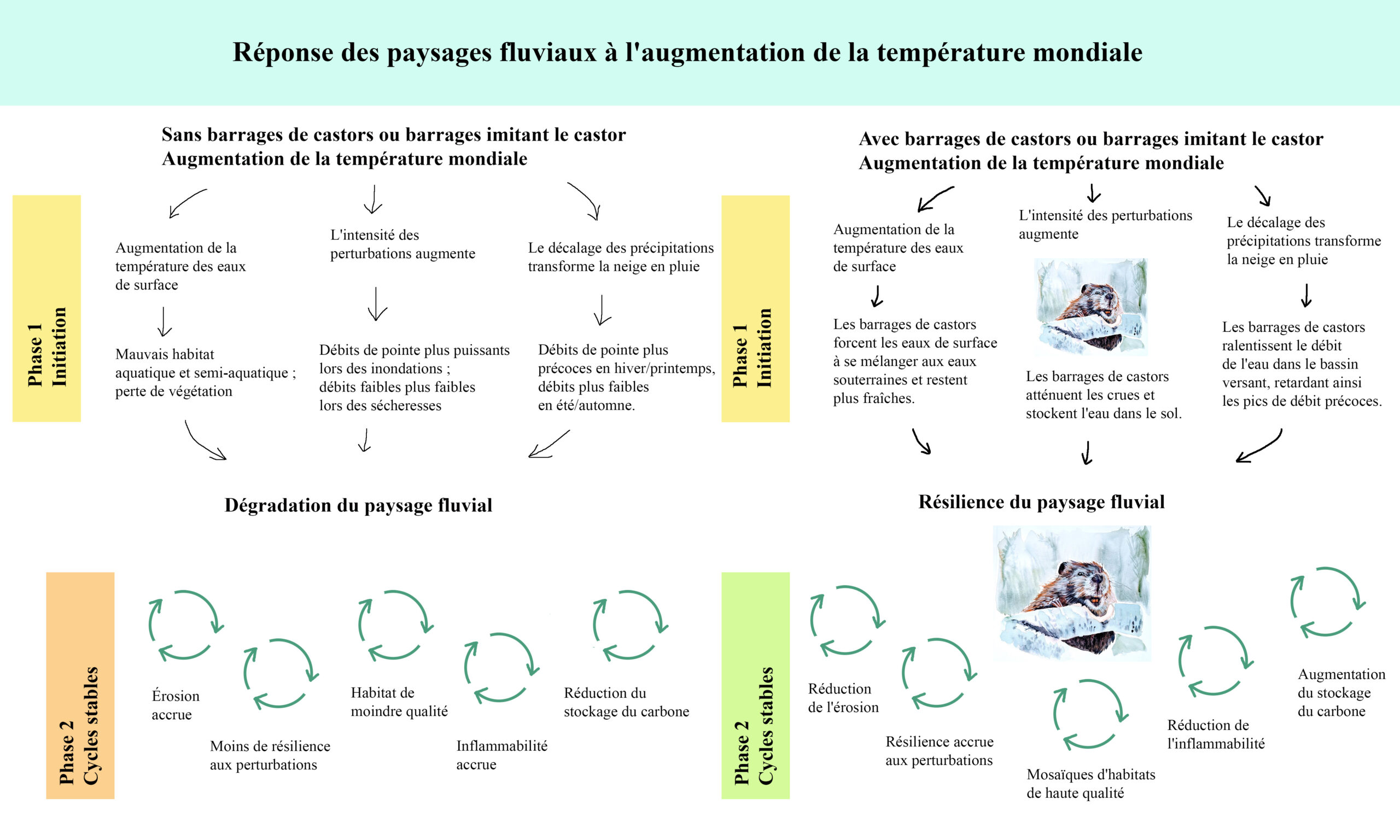
FIGURE 1 : Comparaison des cycles de rétroaction des paysages fluviaux avec l’augmentation de la température mondiale. La phase 1 indique les processus qui sont initiés par le réchauffement des températures mondiales et qui conduisent soit à la dégradation, soit à la résilience. La phase 2 indique les processus qui se produisent une fois que les paysages fluviaux ont déjà atteint un état dégradé ou résilient. Gauche : Cycle de dégradation croissante des paysages fluviaux sans castor ni ouvrages inspirés du castor. A droite : Cycle du maintien de la résilience du paysage fluvial qui peut être atteint en s’associant au castor et en imitant les barrages de castor.
3 – Les interconnexions biologiques, hydrologiques et géomorphologiques de la résilience des plaines alluviales.
Les bassins versants sont généralement décrits par les aspects physiques (par exemple, la direction et le débit de l’écoulement, la superficie, le gradient, les précipitations, la géologie) qui sont censés déterminer toutes les propriétés caractéristiques (Kasprak et al., 2016). Cependant, les bassins versants sont bien plus que la somme de leurs propriétés physiques (Fausch et al., 2002). Les bassins versants, en tant que paysages fluviaux, sont des systèmes bio-géomorphiques dynamiques et riches en énergie. Dans les zones de climat mésique tempéré, les paysages fluviaux ont jusqu’à six ordres de grandeur d’énergie potentielle (chimique) stockée dans la matière organique (à la fois vivante et dans le cycle de décomposition) que l’énergie potentielle (physique) de l’écoulement dans le lit mineur des cours d’eau (Phillips, 2016). Les plaines alluviales connectées sont plus productives que les plaines alluviales déconnectées, en partie en raison de leur capacité à retenir et à extraire l’énergie potentielle chimique des composants biotiques (organiques) du bassin versant (Puttock et al., 2018 ; Wegener et al., 2017). Les plaines alluviales fonctionnelles sont connectées parce que les structures longitudinales augmentent la résistance au mouvement des eaux de surface, forcent l’eau à déborder dans la plaine alluviale et forment une diversité de voies d’écoulement à travers tout le fond de la vallée (Pollock et al., 2014 ; Wheaton et al., 2019). Cette rugosité hydraulique dans le chenal et à travers la plaine alluviale dissipe l’énergie de l’écoulement, ce qui maintient l’équilibre transport-dépôt plutôt du côté des dépôts, mais surtout, elle augmente le temps de séjour de l’eau de surface et de l’eau de l’aquifère hyporhéique, ce qui augmente le cyclage des nutriments et la productivité des écosystèmes (Briggs et al., 2013 ; Helton et al., 2014). Les plaines alluviales connectées aux cours d’eau sont hydrologiquement inefficaces, une caractéristique nécessaire mais souvent sous-estimée.
De nombreux cours d’eau et rivières altérés, ainsi que leurs plaines alluviales, sont dans un état physiquement stable, mais simplifié et dégradé (Cluer & Thorne, 2014). Les systèmes de plaines alluviales connectées sont dynamiques (Naiman et al., 2010). Ils sont dans un état de quasi-équilibre entre de nombreux processus de forçage et sont donc intrinsèquement plus résilients aux perturbations que les cours d’eau dégradés (Silverman et al., 2019 ; Wohl, 2021a ; Wohl et al., 2017, 2021). Mais maintenir l’état de quasi-équilibre d’une plaine alluviale connectée nécessite un apport énergétique continu. Cette situation est similaire à de nombreux systèmes biogéomorphiques marins côtiers bien étudiés – y compris les récifs coralliens côtiers, les marais de mangrove et les marais salants – des écosystèmes mus par la dynamique des vagues, les courants de marée et l’écoulement de l’eau douce, mais contraints par leurs composants biologiques (Alongi, 2008 ; Dame & Patten, 1981 ; Johnson et al., 2019 ; Pethick, 1992).
Dans les systèmes bio-géomorphologiques, les plantes et les animaux forment des structures qui modifient l’environnement physique, ce qui donne lieu à un écosystème plus productif (biomasse générée par unité de surface par unité de temps) que le même endroit ne le serait sans la structure (Viles, 1988 ; C. Wang et al., 2020). Cependant, une fois qu’un système bio-géomorphologique a été dégradé, il ne faut pas grand-chose pour qu’il reste dans cet état (Castro et Thorne, 2019). La restauration de systèmes simplifiés à haute énergie nécessite de rétablir le contrôle par les forces biologiques du milieu géomorphologique (Johnson et al., 2019). Dans les plaines alluviales, bon nombre des plus importantes sources d’énergie externe sont des apports biologiques, notamment le dépôt de matières organiques, la croissance de la végétation et la construction de barrages de castors. Par conséquent, l’adoption d’ingénieurs de l’écosystème comme le castor est la voie rapide vers une connectivité durable du paysage fluvial à faible coût et à fort impact (Brazier et al., 2021 ; Dittbrenner et al., 2018 ; Johnson et al., 2019 ; Pollock et al., 2007, 2014).
4 – Avantages des rivières connectées aux plaines alluviales dans un monde qui se réchauffe
La restauration et la reconnexion des plaines alluviales présentent clairement une multitude d’avantages. Une vallée connectée à une plaine alluviale est intrinsèquement plus diversifiée et productive, non seulement pour les espèces aquatiques, mais aussi sur l’ensemble de la plaine alluviale (Bellmore & Baxter, 2014). Sur la surface de la plaine alluviale humide en saison, la productivité de la végétation et la richesse et la diversité des espèces végétales et animales sont plus élevées que sur une terrasse déconnectée et sèche en permanence (Stella et al., 2011). Dans les chenaux d’un tronçon de plaine alluviale connectée, la productivité primaire est plus élevée, les communautés de macroinvertébrés sont plus riches et plus productives (Nummi et al., 2021 ; Robinson et al., 2020), et la productivité des amphibiens et des poissons est plus élevée (Anderson et al., 2015 ; Bouwes et al., 2016 ; Dauwalter & Walrath, 2018 ; Romansic et al., 2021 ; Wathen et al., 2019) que dans les chenaux simples d’un tronçon déconnecté. Mais, si ces avantages internes sont indépendamment précieux, ils ne représentent qu’une petite fraction des avantages potentiels que les paysages fluviaux restaurés peuvent fournir face au changement climatique. Lorsque nous reconnectons les cours d’eau et les rivières à leurs plaines alluviales, nous effectuons à la fois un travail d’atténuation du changement climatique (ralentir/arrêter la trajectoire du réchauffement climatique) et un travail d’adaptation au changement climatique (renforcer la résilience et la résistance aux perturbations d’origine climatique qui se produisent déjà ; voir le tableau 1).
TABLEAU 1. Résumé schématique de la manière dont les paysages fluviaux de plaine alluviale connectés et déconnectés répondent généralement à plusieurs aspects clés du changement climatique (avec des références sélectionnées abrégées).
Plaines alluviales déconnectées | Plaines alluviales connectées | Références sélectionnées abrégées | |
Température de l’eau (adaptation) | Homogène, plus chaude | Hétérogène, plus froide | Majerova et al., 2015, Weber et al., 2017, Dauwalter & Walrath, 2018 ; Lowry, 1993, Romansic et al., 2021 |
Carbone (atténuation) | Faible potentiel de séquestration | Potentiel de séquestration plus élevé | Wohl, 2013, Laurel & Wohl, 2019 |
Inondations (adaptation) | Faible capacité d’adaptation aux ondes de crue, taux d’érosion plus élevés sur les berges des chenaux en raison de débits de pointe plus puissants. | Grande capacité à accueillir les vagues de crue, érosion moindre des berges du canal due à la dissipation des débits de pointe | Westbrook et al., 2006, Westbrook et al., 2020, Puttock et al., 2021 |
Sécheresses (adaptation) | Faible capacité à maintenir la productivité primaire pendant les périodes sèches prolongées | Forte capacité à maintenir la productivité primaire pendant les périodes sèches prolongées | G. A. Hood & Bayley, 2008, Fairfax & Small, 2018, Dittbrenner et al., 2018 |
Incendies (adaptation) | Inflammabilité plus élevée du combustible. La perte de la végétation riveraine entraîne l’entrée dans la rivière d’intenses débris post-incendie provenant des zones environnantes Inflammabilité plus faible du combustible. | Une végétation riveraine intacte ralentit la pénétration des débris dans la rivière depuis les zones environnantes. Les structures dans le cours d’eau retiennent les sédiments et s’aggradent dans le chenal, inversant l’incision antérieure du canal | Fairfax & Whittle, 2020, Wohl et al, 2022, Weirich, 2021, Whipple, 2019 |
4.1 – Résilience des eaux lentes face aux sécheresses, aux inondations et aux incendies
La diversité des temps de résidence de l’eau dans un réseau fluvial renforce la capacité du paysage fluvial à atténuer les débits de crue pendant les périodes humides et à libérer l’eau stockée sous forme de débit de base pendant les périodes sèches, ce qui permet d’atténuer simultanément les effets de la sécheresse et des inondations (Fairfax et Small, 2018 ; G. A. Hood et Bayley, 2008 ; G. A. Hood et Larson, 2015 ; Puttock et autres, 2021 ; Westbrook et autres, 2006, 2020). Cela permet également de maintenir l’eau dans le sol pendant les périodes de sécheresse prolongée, où elle est accessible à la végétation riveraine (Amlin & Rood, 2003 ; Dittbrenner et al., 2018 ; Fairfax & Small, 2018 ; Puttock et al., 2021 ; Silverman et al., 2019 ; Vivian et al., 2014). Cependant, les paysages fluviaux reliés à la plaine alluviale fonctionnent comme des ralentisseurs de la propagation du feu, car le sol, la végétation et les canaux des cours d’eau sont humides tout au long du parcours et ne brûlent donc pas facilement (Fairfax & Whittle, 2020 ; Weirich, 2021 ; Whipple, 2019 ; Wohl et al., 2022). Par conséquent, de longues étendues de plaines alluviales restaurées pourraient fonctionner comme un réseau de coupe-feu, ralentissant la propagation des feux de forêt et donnant aux humains le temps de contenir les feux de forêt incontrôlés avant qu’ils n’atteignent un état dangereux et hors de contrôle (Fairfax & Whittle, 2020).
4.2 – Biodiversité aquatique stimulée par une eau propre et fraîche
Les paysages fluviaux connectés à la plaine alluviale présentent davantage de gros bois (bois détaché, rondins de bois) à la fois sur le sol et dans le chenal. Les dépôts de bois renforcent en général le forçage physique de la structure des cours d’eau et des plaines alluviales et accroissent le temps de résidence de l’eau et la productivité primaire à la surface de la plaine alluviale (Appling et al., 2014 ; Briggs et al., 2013 ; Collins et al., 2012 ; Helton et al., 2014 ; Magilligan et al., 2008 ; Osei et al., 2015 ; Poole et al., 2008). Mais, la végétation gérée par les castors et la modification hydraulique des cours d’eau (barrages, loges, canaux) fonctionnent de manière similaire, car elles augmentent directement la diversité hydraulique et la productivité de la végétation (Silverman et al., 2019). Ainsi, l’inefficacité hydraulique, quelle qu’en soit la source, entraîne un temps de résidence de l’eau plus long et une augmentation du cycle des nutriments, ce qui améliore la productivité biologique à tous les niveaux trophiques.
Les zones humides, les plaines alluviales inondées, les bassins profonds et d’autres zones d’eau lente dans les paysages fluviaux contribuent à l’évacuation et à la transformation des polluants aquatiques courants tels que les nitrates, les phosphates, les métaux et les excès de sédiments (Klotz, 1998, 2010 ; Kroes & Bason, 2015 ; Maret et al., 1987 ; Muskopf, 2007 ; Puttock et al., 2017, 2018 ; Shepherd & Nairn, 2020, 2021 ; Short et al., 2015). Certains polluants se fixent sur des sédiments fins qui restent en suspension dans la colonne d’eau jusqu’à ce qu’ils atteignent les tronçons à faible vitesse. Une fois que ces sédiments fins sont soit déposés sur la plaine alluviale, soit déposés au fond des étangs et des zones humides, les processus biogéochimiques naturels transforment les nutriments puissants (par exemple, les nitrates) en composés inertes (par exemple, l’azote gazeux) ou facilitent la réabsorption par la végétation aquatique (Yousaf et al., 2021). Cependant, les approches systématiques fondées sur la nature d’atténuation de la pollution à l’échelle du paysage fluvial font défaut, et le rétablissement de l’équilibre biogéochimique naturel des systèmes cours d’eau-zones humides récemment restaurés prend du temps (Weigelhofer et al., 2018). Par conséquent, les structures dans les cours d’eau et les rivières, telles que les ouvrages naturels ou artificiels de castors ou une meilleure connexion des plaines alluviales, peuvent servir de stratégie d’atténuation naturelle à l’échelle du réseau hydrographique. Par exemple, la mise en œuvre d’ouvrage inspirés des castors (BDA), gagne en popularité en tant que stratégie de gestion des terres après un incendie pour atténuer les coulées de débris post-incendie et réduire les cendres et la suie en suspension dans la colonne d’eau (Short et al., 2015).
De nombreuses espèces aquatiques ont des exigences strictes en matière de température de l’eau qui sont régulièrement dépassées à mesure que le climat se réchauffe. Les structures dans les rivières, qu’il s’agisse d’imitations biogéomorphiques construites par l’homme (p. ex. des ouvrages castor ou des amas de bois) ou d’origine naturelle (barrages de castors, débris ligneux), génèrent des gradients de pression hydraulique verticale, forçant une partie de l’écoulement vers le bas à travers le fond de la rivière et dans la zone hyporhéique (Munir et Westbrook, 2020 ; Scamardo et Wohl, 2020 ; Wade et al., 2020). Là, l’eau de surface chaude et l’eau souterraine généralement plus froide peuvent se mélanger avant de retourner dans la rivière en aval (Weber et al., 2017). Le temps de résidence de l’eau sur ces voies d’écoulement varie ; par conséquent, la température de l’eau lorsqu’elle retourne à la rivière varie également. Cependant, le profil thermique très hétérogène du paysage fluvial qui en résulte soutient une variété de vie aquatique ayant des besoins différents en matière de température (Dauwalter & Walrath, 2018 ; Lowry, 1993 ; Majerova et al., 2015, 2020 ; Romansic et al., 2021). Par conséquent, les structures dans le cours d’eau, ou les plaines alluviales connectées, sont une composante essentielle des paysages fluviaux au fonctionnement naturel.
4.3 – Cheminements complexes des eaux – stockage du carbone et mosaïques d’habitats
Les paysages fluviaux complexes d’un point de vue hydrologique offrent une diversité d’habitats entremêlés qui abritent un vaste éventail d’espèces végétales et animales. Les ouvrages de castor naturels sont des composantes particulièrement riches et variées des paysages fluviaux qui hébergent des vitesses d’eau, des températures, des profondeurs, des communautés végétales et des structures géomorphiques très hétérogènes dans des zones relativement petites du paysage fluvial (Larsen et al., 2021 ; Rosell et al., 2005 ; Stringer & Gaywood, 2016). Cette hétérogénéité donne lieu à des habitats particulièrement diversifiés et résilients et explique en grande partie pourquoi les castors sont des espèces clés (Hammerson, 1994 ; Naiman et al., 1986 ; Naiman et al., 1988 ; Pollock et al., 1995). Mais, cette composante bio-fluviale clé des paysages fluviaux est rare car une longue histoire d’impacts anthropiques a simplifié et déconnecté les cours d’eau de leurs plaines alluviales (Fouty, 2018). Par conséquent, la reconnexion des plaines alluviales est souvent invoquée pour améliorer la quantité et la qualité des caractéristiques physiques et biologiques de l’habitat dont ont besoin les poissons, les amphibiens, les oiseaux d’eau et d’autres espèces aquatiques et semi-aquatiques (Anderson et al, 2015 ; Baldwin, 2015 ; Dauwalter & Walrath, 2018 ; W. G. Hood, 2012 ; Kauffman et al., 1997 ; McKinstry et al., 2001 ; Pollock et al., 2004 ; Romansic et al., 2021 ; Snodgrass & Meffe, 1998 ; Wathen et al., 2019 ; Wohl, 2021b ; Wohl et al., 2021).
La complexité des rivières reliées aux plaines alluviales augmente le stockage du carbone par le biais de plusieurs mécanismes, notamment le renforcement de la séquestration dans les forêts riveraines et le dépôt accru de sédiments riches en matières organiques et les dépôts de carbone fibreux dans les environnements périodiquement et régulièrement inondés (Laurel & Wohl, 2019). Mais l’activité de construction d’ouvrages de castors peut augmenter encore plus le stockage de carbone via une réduction supplémentaire de la vitesse d’écoulement des cours d’eau, le trognage régulier des arbres, l’expansion de la superficie des terres périodiquement inondées et l’inondation fréquente de 10 à 30 % du fond de la vallée au débit de base. Même les ouvrages de castors reliques ou inactifs stockent beaucoup plus de carbone que les zones sans présence récente de castors. Des recherches récentes indiquent que les prairies (qui remplacent souvent les forêts riveraines dans les systèmes fluviaux dégradés) stockent en moyenne 40 à 100 tonnes métriques de carbone par hectare, tandis que les complexes de castors actifs et inactifs/récents stockent respectivement 1150 à 1400 et 300 à 400 tonnes métriques de carbone par hectare (Wohl, 2013). Cependant, le bilan carbone complet des plaines alluviales modifiées par les castors est un équilibre complexe dans l’espace et dans le temps. Le carbone enfermé dans la végétation morte sur pied et dans la matière organique déposée dans le lit du cours d’eau et de l’étang est compensé par les émissions de CO2 et de CH4 du cycle de décomposition. Bien que peu documenté, on estime que le bilan net varie de source à puits, en fonction de l’âge de l’étang, de la température et du type de sol et de végétation (Nummi et al., 2018). Ainsi, la restauration de paysages fluviaux, en particulier la restauration de plaines alluviales avec des castors, peut soutenir une augmentation significative du stockage de carbone dans le paysage, et fournir des avantages en matière d’atténuation du climat ainsi que d’adaptation, bien que des questions subsistent concernant les facteurs agissant sur le stockage net de carbone et donc notre capacité à concevoir et à généraliser dans tous les contextes.
4.4 – Dynamique de la plaine alluviale
La végétation de la canopée détermine les niveaux de rayonnement photosynthétique actif (RPA) à la surface des terrasses de la plaine alluviale et dans les canaux des cours d’eau. Les niveaux de RPA, qui dépendent de la disponibilité des nutriments, déterminent les taux de productivité primaire à la surface de la plaine alluviale et dans le chenal du cours d’eau. Ainsi, une canopée forestière riveraine fermée est moins productive qu’un assemblage de plantes de plaine alluviale à plusieurs niveaux et de stature végétale diverse (Ecke et al., 2017). Un assemblage de plantes riveraines mature et de grande stature ne se développe que sur une surface de plaine alluviale stable et peu perturbée qui, à son tour, existe lorsque les taux de migration et de tressage des canaux sont minimes. Cependant, les taux de migration et de tressage du canal sont réduits par les processus qui stabilisent l’emplacement du canal, tels que l’incision et la croissance des racines des plantes profondes et fortes (Hawley & MacMannis, 2019). Ainsi, un assemblage de plantes riveraines maximalement productif est celui qui présente une gamme de hauteur et de structure qui tolère les perturbations dues à la migration et à la formation du canal. Mais le castor est également un agent structurant clé pour les plantes des plaines alluviales (Johnson et al., 2019 ; Westbrook, 2021). La pression de broutage exercée par les castors sélectionne les espèces végétales riveraines qui tolèrent la suppression des branches, des pousses ou des tiges. De nombreuses espèces de plantes adaptées au broutage sont plus productives sous la pression du broutage que sans cette pression. Ainsi, les castors modifient fortement l’hydraulique des cours d’eau et des plaines alluviales en creusant des canaux, des tunnels et des terriers, et en construisant des barrages, des réserves de nourriture et des huttes (G. A. Hood & Larson, 2015).
4.5 – Services écosystémiques
Devrions-nous confier à un gros rongeur des tâches d’ingénierie environnementale aussi critiques ? Si la restauration des paysages fluviaux est vraiment un élément si important de notre plan d’action national pour le climat, ne devrions-nous pas le faire nous-mêmes ? En définitive, l’ampleur des changements à opérer dépasse ce que nous pouvons accomplir et maintenir par nous-mêmes. Cependant, la restauration des paysages fluviaux par les castors présente un retour sur investissement élevé, tant en termes de revenus que de contrôle des dépenses (Baldwin, 2015 ; Blackfeet Nation, 2018 ; Blackfeet Nation & Levitus, 2019 ; Pollock et al., 2015 ; S. Thompson et al., 2021 ; Wheaton et al., 2019). La génération de revenus résulte généralement de l’augmentation du tourisme et des loisirs de plein air (par exemple, la chasse, la pêche, la randonnée, le camping, l’observation de la faune), tandis que la réduction des dépenses provient de la diminution des dépenses en matière d’atténuation des catastrophes naturelles, d‘une meilleure gestion du cycle du carbone, de l’assurance d’une meilleure qualité de l’eau et de conservation de l’eau. Ces services écosystémiques rendus par le castor, ainsi que de nombreux autres qui ne sont pas abordés en détail ici, sont estimés à 69 000 $ par kilomètre carré, par an (S. Thompson et al., 2021). Les avantages économiques secondaires de l’utilisation de la coexistence et du mimétisme du castor dans la restauration des paysages fluviaux contribueraient à compenser le coût déjà faible de la mise en œuvre du mimétisme du castor et de la gestion des conflits entre l’humain et le castor (Boyles et Savitzky, 2009).
4.6 – Surmonter les contraintes institutionnelles de la restauration fondée sur les castors
Nos poissons, nos eaux et nos forêts dépendent de notre volonté d’agir. Nous ne pouvons pas nous contenter de continuer à étudier la situation sans prendre des mesures. Il faut absolument mener plus de recherches pour optimiser et quantifier les impacts de la restauration fondée sur les castors à toutes les échelles spatiales et temporelles. Dans un monde idéal, nous pourrions attendre pour agir que tous les détails soient réglés. Cependant, compte tenu de la trajectoire du changement climatique et des ressources en eau de plus en plus menacées, nous ne disposons tout simplement pas du temps nécessaire pour une telle stratégie. Nous devons donc mettre en œuvre, et continuer à étudier, les méthodes basées sur les processus naturels dans les cours d’eau dégradés du continent, dès maintenant. Nous devrions commencer à réécrire nos politiques de gestion des castors dès aujourd’hui afin de soutenir activement la coexistence plutôt que la gestion létale, de sorte que si et quand les castors arrivent dans un paysage fluvial, ils puissent prospérer. Nous devrions sensibiliser de manière proactive les gestionnaires de la faune, les gestionnaires du territoire et le public à la valeur incroyable que ces ingénieurs de l’écosystème apportent à nos communautés. La science et la pratique peuvent, et doivent, aller de pair.
Il existe certainement des obstacles à la mise en œuvre de la restauration fondée sur les castors. Aux États-Unis, des questions économiques de haut niveau se posent quant à la légalité des droits d’eau sur les cours d’eau où vivent les castors, quant à l’aspect économique des crédits de restauration des cours d’eau et quant à la possibilité de comptabiliser les zones humides créées par les castors. À un niveau socioculturel plus fondamental, les propriétaires fonciers craignent que les castors coupent tous les arbres, inondent les routes, introduisent des maladies hydriques et mangent des poissons menacés d’extinction si on les laisse recoloniser les cours d’eau. Certaines de ces inquiétudes sont fondées sur la réalité. Par exemple, les castors coupent les arbres, mais ils ne les couperont pas tous et il existe des stratégies de gestion non létales, comme le grillage, pour protéger les arbres importants. Cependant, certaines de ces inquiétudes sont fondées sur des mythes, par exemple, les castors sont des herbivores et ne mangent pas de poissons. Comprendre les conflits – réels ou perçus – entre les castors, les humains et les infrastructures humaines est une étape essentielle pour réussir à élaborer et à promouvoir des stratégies de coexistence efficaces (Auster et al., 2019 ; Auster et al., 2021 ; Auster et al., 2022 ; Charnley et al., 2020 ; McKinstry & Anderson, 1999 ; Siemer et al., 2013). Des efforts continus d’éducation et de sensibilisation sont essentiels pour encourager les travaux de restauration en collaboration avec les castors (Morzillo & Needham, 2015).
Des questions subsistent également sur les impacts physiques et écologiques de la restauration fondée sur les castors. Par exemple, on ne sait pas encore très clairement si les impacts de la restauration castor produiront une réponse linéaire ou non linéaire dans le paysage lorsqu’elle est effectuée à des échelles plus grandes que celles examinées précédemment par les chercheurs. Les différentes configurations et constructions d’ouvrages inspirés des castors sont encore expérimentées et comparées entre elles (Davis et al., 2021 ; Munir & Westbrook, 2020), et d’autres recherches sont nécessaires pour déterminer les configurations optimales pour un site spécifique. Il n’y a pas de consensus clair sur la façon de maximiser les chances de succès lors de la relocalisation des castors – comment piéger les castors vivants, combien de temps les mettre en quarantaine, si le site de relocalisation est préparé intentionnellement ou non, si les impacts des castors relocalisés par rapport aux castors in situ diffèrent : toutes ces considérations sont importantes, avec relativement peu de recherches rigoureuses publiées dans la littérature scientifique (Dittbrenner et al., 2018 ; McCreesh et al., 2019 ; McKinstry et al., 2001). Ces inconnues sont valables, mais elles ne doivent pas complètement paralyser les efforts de restauration fondée sur les castors. Tous les cours d’eau n’auront pas ces problèmes, et tous les projets n’ont pas besoin d’atteindre une restauration maximale entièrement optimisée dès la première tentative. De petits gains de restauration valent mieux que l’absence de gains de restauration, et dans une perspective de précaution et d’atténuation des risques, les progrès réalisés graduellement ont une valeur énorme. Il est sage de concentrer les efforts de restauration avec le castor sur les cours d’eau présentant le plus faible risque de conflit avec les humains et le plus fort potentiel de restauration. Il y a tellement de cours d’eau à restaurer qu’il faudra du temps pour mener à bien les projets les plus simples et les plus directs. Mais, au fur et à mesure que les projets progressent, davantage de données peuvent être et seront collectées pour informer les projets futurs. C’est la nature même de la science et de la gestion des terres en général, et appliquer cette philosophie à la restauration castor n’est pas une idée radicale.
5 – Conclusion : nous avons besoin d’ingénieurs (des écosystèmes)
Pour rétablir la pleine fonctionnalité des plaines alluviales reconnectées aux cours d’eau, nous devons reconnaître le rôle essentiel que jouent les composantes biologiques, en particulier le castor. Lorsque nous enlevons les castors des cours d’eau et des rivières, ou que nous les empêchons de se réinstaller dans leurs bassins versants ancestraux, le système cours d’eau-plaine alluviale tombe en ruine (Wohl, 2021b). Une fois qu’ils sont déconnectés de leur plaine alluviale, les cours d’eau encaissés se simplifient en chenaux uniques. Les sédiments et le carbone sont exportés du stockage à long terme, l’eau se réchauffe et devient eutrophisée, le paysage s’assèche et les incendies se propagent sur des kilomètres à travers une étendue uniforme de combustible, le tout laissant peu d’habitat sain pour les poissons et la faune. Mais les plaines alluviales gérées par les castors sont des hauts lieux de la biodiversité, car les étangs et les zones humides des castors servent de puits de carbone, de centres de traitement de l’azote et du phosphore, de réservoirs pour le stockage et le refroidissement de l’eau, et de sites d’atténuation des effets des sécheresses et des inondations. Il est donc impératif de favoriser les zones aménagées par les castors pour les nombreux services qu’elles rendent.
Nous devons mettre en pratique nos connaissances des processus physiques et biologiques des paysages fluviaux fonctionnels et du rôle joué par les castors pour mener une action rapide, globale et durable. Des actions qui résolvent le problème de la dégradation généralisée des ruisseaux, rivières et plaines alluviales d’Amérique du Nord. Des actions qui rétablissent la dynamique naturelle et fonctionnelle des paysages fluviaux afin de permettre des réponses robustes aux perturbations. La restauration des paysages fluviaux, et en particulier la restauration low tech fondée sur les processus et inspirée du castor, devrait être le fondement de notre plan d’action national pour le climat en eau douce.
Remerciements
Cette perspective a grandement bénéficié de la révision et des commentaires de Ben Goldfarb, Nick Bouwes, Martha Otis, Lisa Crozier, Shallin Busch, Alan Puttock et de trois lecteurs anonymes.
Conflits d’intérêts
Les auteurs n’ont déclaré aucun conflit d’intérêt pour cet article.
Contributions des auteurs
Chris E. Jordan : Conceptualisation (à égalité) ; administration du projet (à égalité) ; rédaction – version originale (à égalité) ; rédaction – révision et édition (à égalité). Emily Fairfax : Conceptualisation (à égalité) ; administration du projet (à égalité) ; rédaction – version originale (à égalité) ; rédaction – révision et édition (à égalité).
Références
- Albert, S., & Trimble, T. (2000). Beavers are partners in riparian restoration on the Zuni Indian Reservation. Ecological Restoration, 18(2), 87–92 http://www.jstor.org/stable/43440851
CrossrefGoogle Scholar - Alongi, D. M. (2008). Mangrove forests: Resilience, protection from tsunamis, and responses to global climate change. Estuarine, Coastal and Shelf Science, 76(1), 1–13.
CrossrefWeb of Science®Google Scholar - Amlin, N. M., & Rood, S. B. (2003). Drought stress and recovery of riparian cottonwoods due to water table alteration along Willow Creek, Alberta. Trees, 17, 351–358.
CrossrefWeb of Science®Google Scholar - Anderson, N. L., Paszkowski, C. A., & Hood, G. A. (2015). Linking aquatic and terrestrial environments: Can beaver canals serve as movement corridors for pond-breeding amphibians? Animal Conservation, 18, 287–294.
Wiley Online LibraryWeb of Science®Google Scholar - Appling, A. P., Bernhardt, E. S., & Stanford, J. A. (2014). Floodplain biogeochemical mosaics: A multidimensional view of alluvial soils. Journal of Geophysical Research: Biogeosciences, 119(8), 1538–1553.
Wiley Online LibraryCASWeb of Science®Google Scholar - Auster, R. E., Barr, S. W., & Brazier, R. E. (2021). Improving engagement in managing reintroduction conflicts: Learning from beaver reintroduction. Journal of Environmental Planning and Management, 64(10), 1713–1734. https://doi.org/10.1080/09640568.2020.1837089
CrossrefWeb of Science®Google Scholar - Auster, R. E., Barr, S. W., & Brazier, R. E. (2022). Renewed coexistence: Learning from steering group stakeholders on a beaver reintroduction project in England. European Journal of Wildlife Research, 68, 1. https://doi.org/10.1007/s10344-021-01555-6
CrossrefPubMedWeb of Science®Google Scholar - Auster, R. E., Puttock, A., & Brazier, R. (2019). Unravelling perceptions of Eurasian beaver reintroduction in Great Britain. Area, 52, 364–375. https://doi.org/10.1111/area.12576
Wiley Online LibraryWeb of Science®Google Scholar - Baldwin, J. (2015). Potential mitigation of and adaptation to climate-driven changes in California’s highlands through increased beaver populations. California Fish and Game, 101(4), 218–240.
Web of Science®Google Scholar - Beechie, T. J., Imaki, H., Greene, J., Wade, A. A., Wu, H., Pess, G. R., Roni, P., Kimball, J. S., Stanford, J. A., Kiffney, P. M., & Mantua, N. J. (2013). Restoring salmon habitat for a changing climate. River Research and Applications, 29(8), 939–960. https://doi.org/10.1002/rra.2590
Wiley Online LibraryWeb of Science®Google Scholar - Beechie, T. J., Sear, D. A., Olden, J. D., Pess, G. R., Buffington, J. M., Moir, H., Roni, P., & Pollock, M. M. (2010). Process-based principles for restoring river ecosystems. Bioscience, 60, 209–222.
CrossrefWeb of Science®Google Scholar - Bellmore, J. R., & Baxter, C. V. (2014). Effects of geomorphic process domains on river ecosystems: A comparison of floodplain and confined valley segments. River Research and Applications, 30(5), 617–630.
Wiley Online LibraryWeb of Science®Google Scholar - Blackfeet Nation. (2018). Blackfeet climate change adaptation plan. https://blackfeetclimatechange.com/our-environment/climate-change-adaptation-plan/
Google Scholar - Blackfeet Nation, & Levitus, J. (2019). The Ksik Stakii Project beaver mimicry guidebook. https://bcapwebsite.files.wordpress.com/2019/12/beaver-mimicry-guidebook.pdf
Google Scholar - Bouwes, N., Weber, N., Jordan, C. E., Saunders, W. C., Tattam, I. A., Volk, C., Wheaton, J. M., & Pollock, M. M. (2016). Ecosystem experiment reveals benefits of natural and simulated beaver dams to a threatened population of steelhead (Oncorhynchus mykiss). Scientific Reports, 6, 28581. https://doi.org/10.1038/srep28581
CrossrefCASPubMedWeb of Science®Google Scholar - Boyles, S. L., & Savitzky, B. A. (2009). An analysis of the efficacy and comparative costs of using flow devices to resolve conflicts with North American beavers along roadways in the coastal plain of Virginia. In R. M. Timm & M. B. Madon (Eds.), Proceedings of the 23rd Vertebrate Pest Conference (Vol. 2008, pp. 47–52). University of California, Davis.
Google Scholar - Brazier, R. E., Puttock, A., Graham, H. A., Auster, R. E., Davies, K. H., & Brown, C. M. L. (2021). Beaver: Nature’s ecosystem engineers. WIREs Water, 8(1), e1494 https://www.ncbi.nlm.nih.gov/pubmed/33614026
Wiley Online LibraryPubMedWeb of Science®Google Scholar - Briggs, M. A., Lautz, L. K., Hare, D. K., & González-Pinzón, R. (2013). Relating hyporheic fluxes, residence times, and redox-sensitive biogeochemical processes upstream of beaver dams. Freshwater Science, 32(2), 622–641.
CrossrefWeb of Science®Google Scholar - Burchsted, D., Daniels, M., Thorson, R., & Vokoun, J. (2010). The river discontinuum: Applying beaver modifications to baseline conditions for restoration of forested headwaters. Bioscience, 60(11), 908–922.
CrossrefWeb of Science®Google Scholar - Castro, J. M., & Thorne, C. R. (2019). The stream evolution triangle: Integrating geology, hydrology, and biology. River Research and Applications, 35(4), 315–326.
Wiley Online LibraryWeb of Science®Google Scholar - Charnley, S., Gosnell, H., Davee, R., & Abrams, J. (2020). Ranchers and beavers: Understanding the human dimensions of beaver-related stream restoration on western rangelands. Rangeland Ecology & Management, 73(5), 712–723.
CrossrefWeb of Science®Google Scholar - Ciotti, D. C., McKee, J., Pope, K. L., Kondolf, G. M., & Pollock, M. M. (2021). Design criteria for process-based restoration of fluvial systems. Bioscience, 71(8), 831–845.
CrossrefWeb of Science®Google Scholar - Cluer, B., & Thorne, C. (2014). A stream evolution model integrating habitat and ecosystem benefits. River Research and Applications, 30(2), 135–154.
Wiley Online LibraryWeb of Science®Google Scholar - Collins, B. D., Montgomery, D. R., Fetherston, K. L., & Abbe, T. B. (2012). The floodplain large-wood cycle hypothesis: A mechanism for the physical and biotic structuring of temperate forested alluvial valleys in the North Pacific coastal ecoregion. Geomorphology, 139–140, 460–470.
CrossrefWeb of Science®Google Scholar - Cornell, R., Andronescu, A., & Nguyen, K. (2011). The effects of beaver dams on water quality and habitat. Dept. of Earth and Atmospheric Sciences, Metropolitan State College of Denver.
Google Scholar - Dame, R. F., & Patten, B. C. (1981). Analysis of energy flows in an intertidal oyster reef. Marine Ecology Progress Series, 5(2), 115–124.
CrossrefWeb of Science®Google Scholar - Dauwalter, D. C., & Walrath, J. D. (2018). Beaver dams, streamflow complexity, and the distribution of a rare minnow, Lepidomeda copei. Ecology of Freshwater Fish., 27, 606–616. https://doi.org/10.1111/eff.12374
Wiley Online LibraryWeb of Science®Google Scholar - Davee, R., Gosnell, H., & Charnley, S. (2019). Using beaver dam analogues for fish and wildlife recovery on public and private rangelands in eastern Oregon (Res. Pap. PNW-RP-612; pp. 29). US Department of Agriculture, Forest Service, Pacific Northwest Research Station.
Google Scholar - Davis, J., Lautz, L., Kelleher, C., Vidon, P., Russoniello, C., & Pearce, C. (2021). Evaluating the geomorphic channel response to beaver dam analog installation using unoccupied aerial vehicles. Earth Surface Processes and Landforms, 46(12), 2349–2364.
Wiley Online LibraryWeb of Science®Google Scholar - Dittbrenner, B. J., Pollock, M. M., Schilling, J. W., Olden, J. D., Lawler, J. J., & Torgersen, C. E. (2018). Modeling intrinsic potential for beaver (Castor canadensis) habitat to inform restoration and climate change adaptation. PLoS One, 13(2), e0192538 https://www.ncbi.nlm.nih.gov/pubmed/29489853
CrossrefPubMedWeb of Science®Google Scholar - Ecke, F., Levanoni, O., Audet, J., Carlson, P., Eklöf, K., Hartman, G., McKie, B., Ledesma, J., Segersten, J., Truchy, A., & Futter, M. (2017). Meta-analysis of environmental effects of beaver in relation to artificial dams. Environmental Research Letters, 12(11), 113002.
CrossrefWeb of Science®Google Scholar - Environmental Law Institute (ELI). (2016). Stream mitigation: Science, policy, and practice. Environmental Law Institute https://www.eli.org/sites/default/files/eli-pubs/stream-mitigation-science-policy-and-practice-finalreport.pdf
Google Scholar - Fairfax, E., & Small, E. E. (2018). Using remote sensing to assess the impact of beaver damming on riparian evapotranspiration in an arid landscape. Ecohydrology, 11(7), 1–14. https://doi.org/10.1002/eco.1993
Wiley Online LibraryWeb of Science®Google Scholar - Fairfax, E., & Whittle, A. (2020). Smokey the beaver: Beaver-dammed riparian corridors stay green during wildfire throughout the western United States. Ecological Applications, 30(8), 1–8.
Wiley Online LibraryWeb of Science®Google Scholar - Fausch, K. D., Torgersen, C. E., Baxter, C. V., & Li, H. W. (2002). Landscapes to riverscapes: Bridging the gap between research and conservation of stream fishes: A continuous view of the river is needed to understand how processes interacting among scales set the context for stream fishes and their habitat. Bioscience, 52(6), 483–498. https://doi.org/10.1641/0006-3568(2002)052[0483:LTRBTG]2.0.CO;2
CrossrefWeb of Science®Google Scholar - Feit, H. A. (1986). James Bay Cree Indian Management and Moral Considerations of Fur bearers. In Native people and renewable resource management. 1986 symposium of the Alberta Society of Professional Biologists (ASPB) (pp. 49–65). ASPB.
Google Scholar - Finco, M., Quayle, B., Zhang, Y., Lecker, J., Megown, K. A., & Brewer, C. K. (2012). Monitoring trends and burn severity (MTBS): Monitoring wildfire activity for the past quarter century using Landsat data. Paper presented at the In: Morin, Randall S.; Liknes, Greg C. comps. Moving from status to trends: Forest Inventory and Analysis (FIA) symposium 2012; 2012 December 4–6; Baltimore, MD. Gen. Tech. Rep. NRS-P-105 (pp. 222–228). US Department of Agriculture, Forest Service, Northern Research Station [CD-ROM].
Google Scholar - Foster, C. N., Banks, S. C., Cary, G. J., Johnson, C. N., Lindenmayer, D. B., & Valentine, L. E. (2020). Animals as agents in fire regimes. Trends in Ecology & Evolution, 35(4), 346–356.
CrossrefPubMedWeb of Science®Google Scholar - Fouty, S. C. (2018). Euro-American beaver trapping and its long-term impact on drainage network form and function, water abundance, delivery, and system stability. In Johnson, R., Carothers, S. W., Finch, D. M., Kingsley, K. J., Stanley, J. T. (Tech. eds.). Riparian research and management: Past, present, future: Volume 1. Gen. Tech. Rep. RMRS-GTR-377. USDA USFS Rocky Mountain Research Station (pp. 226). https://doi.org/10.2737/RMRS-GTR-377.
Google Scholar - Gadgil, M., Berkes, F., & Folke, C. (1993). Indigenous knowledge for biodiversity conservation. Ambio, 22(2), 151–156.
Web of Science®Google Scholar - Goss, M., Swain, D. L., Abatzoglou, J. T., Sarhadi, A., Kolden, C. A., Williams, A. P., & Diffenbaugh, N. S. (2020). Climate change is increasing the likelihood of extreme autumn wildfire conditions across California. Environmental Research Letters, 15(9), 094016.
CrossrefWeb of Science®Google Scholar - Hammerson, G. A. (1994). Beaver (Castor canadensis): Ecosystem alterations, management, and monitoring. Natural Areas Journal, 14(1), 44–57.
Web of Science®Google Scholar - Hawley, R. J., & MacMannis, K. R. (2019). Tree roots as a dominant agent of streambed habitat, profile pattern, and grade control. Geomorphology, 343, 81–91.
CrossrefWeb of Science®Google Scholar - Helton, A. M., Poole, G. C., Payn, R. A., Izurieta, C., & Stanford, J. A. (2014). Relative influences of the river channel, floodplain surface, and alluvial aquifer on simulated hydrologic residence time in a montane river floodplain. Geomorphology, 205, 17–26 https://www.sciencedirect.com/science/article/pii/S0169555X12000189
CrossrefWeb of Science®Google Scholar - Hood, G. A., & Bayley, S. E. (2008). Beaver (Castor canadensis) mitigate the effects of climate on the area of open water in boreal wetlands in western Canada. Biological Conservation, 141(2), 556–567.
CrossrefWeb of Science®Google Scholar - Hood, G. A., & Larson, D. G. (2015). Ecological engineering and aquatic connectivity: A new perspective from beaver-modified wetlands. Freshwater Biology, 60(1), 198–208.
Wiley Online LibraryWeb of Science®Google Scholar - Hood, W. G. (2012). Beaver in tidal marshes: Dam effects on low-tide channel pools and fish use of estuarine habitat. Wetlands, 32(3), 401–410.
CrossrefWeb of Science®Google Scholar - Ives, R. L. (1942). The beaver-meadow complex. Journal of Geomorphology, 5(3), 191–203.
Google Scholar - Janzen, K., & Westbrook, C. J. (2011). Hyporheic flows along a channelled peatland: Influence of beaver dams. Canadian Water Resources Journal, 36(4), 331–347.
CrossrefWeb of Science®Google Scholar - Johnson, M. F., Thorne, C. R., Castro, J. M., Kondolf, G. M., Mazzacano, C. S., Rood, S. B., & Westbrook, C. (2019). Biomic river restoration: A new focus for river management. River Research and Applications, 36(1), 3–12.
Wiley Online LibraryWeb of Science®Google Scholar - Kasprak, A., Hough-Snee, N., Beechie, T., Bouwes, N., Brierley, G., Camp, R., Fryirs, K., Imaki, H., Jensen, M., O’Brien, G., Rosgen, D., & Wheaton, J. (2016). The blurred line between form and process: A comparison of stream channel classification frameworks. PLoS One, 11(3), e0150293.
CrossrefPubMedWeb of Science®Google Scholar - Kauffman, J. B., Beschta, R. L., Otting, N., & Lytjen, D. (1997). An ecological perspective of riparian and stream restoration in the Western United States. Fisheries, 22(5), 12–24.
Wiley Online LibraryWeb of Science®Google Scholar - Keeble-Toll, A. K. (2018). Braiding pine: Weaving traditional knowledge, community need, and scientific method in Sierra Nevada meadow restoration. https://ui.adsabs.harvard.edu/abs/2018AGUFMPA51C0784K
Google Scholar - Kimmerer, R. W. (2000). Native knowledge for native ecosystems. Journal of Forestry, 98(8), 4–9.
Web of Science®Google Scholar - Kimmerer, R. W., & Lake, F. K. (2001). The role of indigenous burning in land management. Journal of Forestry, 99(11), 36–41.
Web of Science®Google Scholar - Klotz, R. L. (1998). Influence of beaver ponds on the phosphorus concentration of stream water. Canadian Journal of Fisheries and Aquatic Sciences, 55, 1228–1235.
CrossrefCASWeb of Science®Google Scholar - Klotz, R. L. (2010). Reduction of high nitrate concentrations in a Central New York state stream impounded by beaver. Northeastern Naturalist, 17(3), 349–356.
CrossrefWeb of Science®Google Scholar - Kroes, D. E., & Bason, C. W. (2015). Sediment-trapping by beaver ponds in streams of the mid-Atlantic Piedmont and coastal plain, USA. Southeastern Naturalist, 14(3), 577–595.
CrossrefWeb of Science®Google Scholar - Larsen, A., Larsen, J. R., & Lane, S. N. (2021). Dam builders and their works: Beaver influences on the structure and function of river corridor hydrology, geomorphology, biogeochemistry and ecosystems. Earth-Science Reviews, 218, 103623.
CrossrefCASWeb of Science®Google Scholar - Laurel, D., & Wohl, E. (2019). The persistence of beaver-induced geomorphic heterogeneity and organic carbon stock in river corridors. Earth Surface Processes and Landforms, 44(1), 342–353.
Wiley Online LibraryWeb of Science®Google Scholar - Lazar, J. G., Addy, K., Gold, A. J., Groffman, P. M., McKinney, R. A., & Kellogg, D. Q. (2015). Beaver ponds: Resurgent nitrogen sinks for rural watersheds in the northeastern United States. Journal of Environmental Quality, 44(5), 1684–1693.
Wiley Online LibraryCASPubMedWeb of Science®Google Scholar - Lowry, M. M. (1993). Groundwater elevations and temperature adjacent to a beaver pond in central Oregon (Thesis). Dept. of Forest Engineering, Oregon State University.
Google Scholar - Macfarlane, W. W., Wheaton, J. M., Bouwes, N., Jensen, M. L., Gilbert, J. T., Hough-Snee, N., & Shivik, J. A. (2015). Modeling the capacity of riverscapes to support beaver dams. Geomorphology, 277, 72–99.
CrossrefWeb of Science®Google Scholar - Magilligan, F. J., Nislow, K. H., Fisher, G. B., Wright, J., Mackey, G., & Laser, M. (2008). The geomorphic function and characteristics of large woody debris in low gradient rivers, coastal Maine, USA. Geomorphology, 97(3–4), 467–482.
CrossrefWeb of Science®Google Scholar - Majerova, M., Neilson, B. T., Schmadel, N. M., Wheaton, J. M., & Snow, C. J. (2015). Impacts of beaver dams on hydrologic and temperature regimes in a mountain stream. Hydrology and Earth System Sciences Discussions, 12(1), 839–878.
CrossrefGoogle Scholar - Majerova, M., Neison, B. T., & Roper, B. B. (2020). Beaver dam influences on streamflow hydraulic properties and thermal regimes. Science of the Total Environment, 718(134853), 1–14. https://www.sciencedirect.com/science/article/pii/S0048969719348454
Google Scholar - Maret, T. J., Parker, M., & Fannin, T. E. (1987). The effect of beaver ponds on the nonpoint source water quality of a stream in southwestern Wyoming. Water Research, 21(3), 263–268.
CrossrefCASWeb of Science®Google Scholar - McCreesh, R. K., Fox-Dobbs, K., Wimberger, P., Woodruff, K., Holtgrieve, G., & Pool, T. K. (2019). Reintroduced beavers rapidly influence the storage and biogeochemistry of sediments in headwater streams (Methow River, Washington). Northwest Science, 93(2), 112–121.
CrossrefWeb of Science®Google Scholar - McKinstry, M. C., & Anderson, S. H. (1999). Attitudes of private-and public-land managers in Wyoming, USA, toward beaver. Environmental Management, 23(1), 95–101.
CASPubMedWeb of Science®Google Scholar - McKinstry, M. C., Caffrey, P., & Anderson, S. H. (2001). The importance of beaver to wetland habitats and waterfowl in Wyoming. Journal of the American Water Resources Association, 37(6), 1571–1577.
Wiley Online LibraryWeb of Science®Google Scholar - Morgan, L. H. (1868). The American beaver and his works. JB Lippincott.
CrossrefGoogle Scholar - Mori, A. S., & Johnson, E. A. (2013). Assessing possible shifts in wildfire regimes under a changing climate in mountainous landscapes. Forest Ecology and Management, 310, 875–886.
CrossrefWeb of Science®Google Scholar - Morzillo, A. T., & Needham, M. D. (2015). Landowner incentives and normative tolerances for managing beaver impacts. Human Dimensions of Wildlife, 20(6), 514–530. https://doi.org/10.1080/10871209.2015.1083062
CrossrefWeb of Science®Google Scholar - Munir, T. M., & Westbrook, C. J. (2020). Beaver dam analogue configurations influence stream and riparian water table dynamics of a degraded spring-fed creek in the Canadian Rockies. River Research and Applications, 37(3), 330–342.
Wiley Online LibraryWeb of Science®Google Scholar - Muskopf, S. A. (2007). The effect of beaver (Castor canadensis) dam removal on total phosphorus concentration in Taylor Creek and wetland, South Lake Tahoe. Humboldt State University.
Google Scholar - Naiman, R. J., Decamps, H., & McClain, M. E. (2010). Riparia: Ecology, conservation, and management of streamside communities. Elsevier.
Google Scholar - Naiman, R. J., Johnston, C. A., & Kelley, J. C. (1988). Alteration of North American streams by beaver. Bioscience, 38(11), 753–762.
CrossrefWeb of Science®Google Scholar - Naiman, R. J., Melillo, J. M., & Hobbie, J. E. (1986). Ecosystem alteration of boreal forest streams by beaver (Castor canadensis). Ecology, 67(5), 1254–1269.
Wiley Online LibraryWeb of Science®Google Scholar - Nash, C. S., Grant, G. E., Charnley, S., Dunham, J. B., Gosnell, H., Hausner, M. B., Pilliod, D. S., & Taylor, J. D. (2021). Great expectations: Deconstructing the process pathways underlying beaver-related restoration. Bioscience, 71(3), 249–267. https://doi.org/10.1093/biosci/biaa165
CrossrefWeb of Science®Google Scholar - Nash, C. S., Selker, J. S., Grant, G. E., Lewis, S. L., & Noël, P. (2018). A physical framework for evaluating net effects of wet meadow restoration on late-summer streamflow. Ecohydrology, 11(5), e1953.
Wiley Online LibraryWeb of Science®Google Scholar - Neff, D. J. (1957). Ecological effects of beaver habitat abandonment in the Colorado Rockies. The Journal of Wildlife Management, 21(1), 80–84.
CrossrefGoogle Scholar - Nummi, P., Liao, W., van der Schoor, J., & Loehr, J. (2021). Beaver creates early successional hotspots for water beetles. Biodiversity and Conservation, 30(10), 2655–2670.
CrossrefWeb of Science®Google Scholar - Nummi, P., Vehkaoja, M., Pumpanen, J., & Ojala, A. (2018). Beavers affect carbon biogechemistry: Both short-term and long-term processes are involved. Mammal Review, 48(4), 298–311. https://doi.org/10.1111/mam.12134
Wiley Online LibraryWeb of Science®Google Scholar - Osei, N. A., Gurnell, A. M., & Harvey, G. L. (2015). The role of large wood in retaining fine sediment, organic matter and plant propagules in a small, single-thread forest river. Geomorphology, 235, 77–87.
CrossrefWeb of Science®Google Scholar - Pearce, C., Vidon, P., Lautz, L., Kelleher, C., & Davis, J. (2021a). Impact of beaver dam analogues on hydrology in a semi-arid floodplain. Hydrological Processes, 35(7), e14275. https://doi.org/10.1002/hyp.14275
Wiley Online LibraryWeb of Science®Google Scholar - Pearce, C., Vidon, P., Lautz, L., Kelleher, C., & Davis, J. (2021b). Short-term impact of beaver dam analogues on streambank erosion and deposition in semi-arid landscapes of the western USA. River Research and Applications, 37(7), 1032–1037.
Wiley Online LibraryWeb of Science®Google Scholar - Pethick, J. (1992). Saltmarsh geomorphology. In Saltmarshes: Morphodynamics, conservation and engineering significance (pp. 41–62). Cambridge, England: Cambridge University Press.
Google Scholar - Phillips, J. D. (2016). Landforms as extended composite phenotypes. Earth Surface Processes and Landforms, 41(1), 16–26.
Wiley Online LibraryWeb of Science®Google Scholar - Pilliod, D. S., Rohde, A. T., Charnley, S., Davee, R. R., Dunham, J. B., Gosnell, H., Grant, G. E., Hausner, M. B., Huntington, J. L., & Nash, C. (2017). Survey of beaver-related restoration practices in rangeland streams of the Western USA. Environmental Management, 61(1), 58–68. https://doi.org/10.1007/s00267-017-0957-6
CrossrefPubMedWeb of Science®Google Scholar - Pollock, M. M., Beechie, T. J., & Jordan, C. E. (2007). Geomorphic changes upstream of beaver dams in Bridge Creek, an incised stream channel in the interior Columbia River basin, eastern Oregon. Earth Surface Processes and Landforms, 32(8), 1174–1185.
Wiley Online LibraryWeb of Science®Google Scholar - Pollock, M. M., Beechie, T. J., Wheaton, J. M., Jordan, C. E., Bouwes, N., Weber, N., & Volk, C. (2014). Using beaver dams to restore incised stream ecosystems. Bioscience, 64(4), 279–290.
CrossrefWeb of Science®Google Scholar - Pollock, M. M., Castro, J., Jordan, C. E., Lewallen, G., & Woodruff, K. (2015). The beaver restoration guidebook: Working with beaver to restore streams, wetlands, and floodplains (Version 1.02) (p. 189). United States Fish and Wildlife Service.
Google Scholar - Pollock, M. M., Naiman, R. J., Erickson, H. E., Johnston, C. A., Pastor, J., & Pinay, G. (1995). Beaver as engineers—Influences on biotic and abiotic characteristics of drainage basins. Linking Species & Ecosystems, 117–126. Boston, MA: Springer. https://doi.org/10.1007/978-1-4615-1773-3
CrossrefGoogle Scholar - Pollock, M. M., Pess, G., & Beechie, T. J. (2004). The importance of beaver ponds to Coho Salmon production in the Stillaguamish River basin, Washington, USA. North American Journal of Fisheries Management, 24, 749–760.
Wiley Online LibraryWeb of Science®Google Scholar - Poole, G. C., O’Daniel, S. J., Jones, K. L., Woessner, W. W., Bernhardt, E. S., Helton, A. M., Stanford, J. A., Boer, B. R., & Beechie, T. J. (2008). Hydrologic spiralling: The role of multiple interactive flow paths in stream ecosystems. River Research and Applications, 24(7), 1018–1031. https://doi.org/10.1002/rra.1099
Wiley Online LibraryWeb of Science®Google Scholar - Puttock, A., Graham, H. A., Ashe, J., Luscombe, D. J., & Brazier, R. E. (2021). Beaver dams attenuate flow: A multi-site study. Hydrological Processes, 35(2), e14017 https://www.ncbi.nlm.nih.gov/pubmed/33678948
Wiley Online LibraryPubMedWeb of Science®Google Scholar - Puttock, A., Graham, H. A., Carless, D., & Brazier, R. E. (2018). Sediment and nutrient storage in a beaver engineered wetland. Earth Surface Processes and Landforms, 43, 2358–2370. https://doi.org/10.1002/esp.4398
Wiley Online LibraryCASPubMedWeb of Science®Google Scholar - Puttock, A., Graham, H. A., Cunliffe, A. M., Elliott, M., & Brazier, R. E. (2017). Eurasian beaver activity increases water storage, attenuates flow and mitigates diffuse pollution from intensively-managed grasslands. Science of the Total Environment, 576, 430–443 https://www.ncbi.nlm.nih.gov/pubmed/27792958
CrossrefCASPubMedWeb of Science®Google Scholar - Robinson, C. T., Schweizer, P., Larsen, A., Schubert, C. J., & Siebers, A. R. (2020). Beaver effects on macroinvertebrate assemblages in two streams with contrasting morphology. Science of the Total Environment, 722, 137899.
CrossrefCASPubMedWeb of Science®Google Scholar - Romansic, J. M., Nelson, N. L., Moffett, K. B., & Piovia-Scott, J. (2021). Beaver dams are associated with enhanced amphibian diversity via lengthened hydroperiods and increased representation of slow-developing species. Freshwater Biology, 66(3), 481–494. https://doi.org/10.1111/fwb.13654
Wiley Online LibraryWeb of Science®Google Scholar - Rosell, F., Bozser, O., Collen, P., & Parker, H. (2005). Ecological impact of beavers Castor fiber and Castor canadensis and their ability to modify ecosystems. Mammal Review, 35(3), 248–276.
Wiley Online LibraryWeb of Science®Google Scholar - Ruedemann, R., & Schoonmaker, W. J. (1938). Beaver dams as geologic agents. Science, 88(2292), 523–525.
CrossrefCASPubMedGoogle Scholar - Sands, A., & Howe, G. (1977). Riparian forests in California, their ecology and conservation. Paper presented at the Importance, Preservation and Management of Riparian Habitat: A Symposium, Tuscon, AZ.
Google Scholar - Scamardo, J., & Wohl, E. (2020). Sediment storage and shallow groundwater response to beaver dam analogues in the Colorado front range, USA. River Research and Applications, 36(3), 398–409.
Wiley Online LibraryWeb of Science®Google Scholar - Seton, E. T. (1929). Lives of game animals (Vol. 4, pp. 441–501). Doubleday, Doran and Co.
Google Scholar - Shepherd, N. L., & Nairn, R. W. (2020). Metals retention in a net alkaline mine drainage impacted stream due to the colonization of the North American beaver (Castor canadensis). Science of the Total Environment, 731, 139203.
CrossrefCASPubMedWeb of Science®Google Scholar - Shepherd, N. L., & Nairn, R. W. (2021). Induced mobilization of stored metal precipitates from beaver (Castor canadensis) created wetlands on a mine drainage impacted stream. Wetlands Ecology and Management, 30, 127–137.
CrossrefWeb of Science®Google Scholar - Sherriff, L. (2021). Beaver believers: Native Americans promote resurgence of ‘nature’s engineers’. The Guardian.
Google Scholar - Short, L. E., Gabet, E. J., & Hoffman, D. F. (2015). The role of large woody debris in modulating the dispersal of a post-fire sediment pulse. Geomorphology, 246, 351–358.
CrossrefWeb of Science®Google Scholar - Siemer, W. F., Jonker, S. A., Decker, D. J., & Organ, J. F. (2013). Toward an understanding of beaver management as human and beaver densities increase. Human-Wildlife Interactions, 7(1), 114–131.
Google Scholar - Silverman, N. L., Allred, B. W., Donnelly, J. P., Chapman, T. B., Maestas, J. D., Wheaton, J. M., & White, J. (2019). Low-tech riparian and wet meadow restoration increases vegetation productivity and resilience across semiarid rangelands. Restoration Ecology, 27(2), 269–278. https://doi.org/10.1111/rec.12869
Wiley Online LibraryWeb of Science®Google Scholar - Skidmore, P., & Wheaton, J. M. (2022). Natural infrastructure—Can restored riverscapes help us adapt to climate change. Anthropocene. https://doi.org/10.13140/RG.2.2.33525.86248
CrossrefGoogle Scholar - Snodgrass, J. W., & Meffe, G. K. (1998). Influence of beavers on stream fish assemblages: Effects of pond age and watershed position. Ecology, 79(3), 928–942.
Wiley Online LibraryWeb of Science®Google Scholar - Stella, J. C., Hayden, M. K., Battles, J. J., Piégay, H., Dufour, S., & Fremier, A. K. (2011). The role of abandoned channels as refugia for sustaining pioneer riparian forest ecosystems. Ecosystems, 14(5), 776–790.
CrossrefWeb of Science®Google Scholar - Stott, P. (2016). How climate change affects extreme weather events. Science, 352(6293), 1517–1518.
CrossrefCASPubMedWeb of Science®Google Scholar - Stringer, A. P., & Gaywood, M. J. (2016). The impacts of beavers Castor spp. on biodiversity and the ecological basis for their reintroduction to Scotland, UK. Mammal Review, 46(4), 270–283.
Wiley Online LibraryWeb of Science®Google Scholar - Swain, D. L. (2021). A shorter, sharper rainy season amplifies California wildfire risk. Geophysical Research Letters, 48(5), 1–5.
Wiley Online LibraryWeb of Science®Google Scholar - Thompson, D. M., & Stull, G. N. (2002). The development and historic use of habitat structures in channel restoration in the United States: The grand experiment in fisheries management. Géographie Physique et Quaternaire, 56(1), 45–60.
CrossrefGoogle Scholar - Thompson, S., Vehkaoja, M., Pellikka, J., & Nummi, P. (2021). Ecosystem services provided by beavers Castor spp. Mammal Review, 51(1), 25–39.
Wiley Online LibraryWeb of Science®Google Scholar - USEPA. (2013). National rivers and streams assessment 2013–2014: A collaborative survey (EPA-841-R-19-001).
Google Scholar - Viles, H. A. (1988). Biogeomorphology: B. Blackwell.
Google Scholar - Vivian, L. M., Godfree, R. C., Colloff, M. J., Mayence, C. E., & Marshall, D. J. (2014). Wetland plant growth under contrasting water regimes associated with river regulation and drought: Implications for environmental water management. Plant Ecology, 215(9), 997–1011.
CrossrefWeb of Science®Google Scholar - Wade, J., Lautz, L., Kelleher, C., Vidon, P., Davis, J., Beltran, J., & Pearce, C. (2020). Beaver dam analogues drive heterogeneous groundwater–surface water interactions. Hydrological Processes, 34(26), 5340–5353.
Wiley Online LibraryWeb of Science®Google Scholar - Wang, C., Smolders, S., Callaghan, D. P., van Belzen, J., Bouma, T. J., Hu, Z., Wen, Q., & Temmerman, S. (2020). Identifying hydro-geomorphological conditions for state shifts from bare tidal flats to vegetated tidal marshes. Remote Sensing, 12(14), 2316. https://doi.org/10.3390/rs12142316
CrossrefWeb of Science®Google Scholar - Wang, X., Shaw, E. L., Westbrook, C. J., & Bedard-Haughn, A. (2018). Beaver dams induce hyporheic and biogeochemical changes in riparian areas in a mountain peatland. Wetlands, 38(5), 1017–1032.
CrossrefWeb of Science®Google Scholar - Wathen, G., Allgeier, J. E., Bouwes, N., Pollock, M. M., Schindler, D. E., & Jordan, C. E. (2019). Beaver activity increases habitat complexity and spatial partitioning by steelhead trout. Canadian Journal of Fisheries and Aquatic Sciences, 76(7), 1086–1095.
CrossrefWeb of Science®Google Scholar - Weber, N., Bouwes, N., Pollock, M. M., Volk, C., Wheaton, J. M., Wathen, G., Wirtz, J., & Jordan, C. E. (2017). Alteration of stream temperature by natural and artificial beaver dams. PLoS One, 12(5), e0176313 https://www.ncbi.nlm.nih.gov/pubmed/28520714
CrossrefPubMedWeb of Science®Google Scholar - Wegener, P., Covino, T., & Wohl, E. E. (2017). Beaver-mediated lateral hydrologic connectivity, fluvial carbon and nutrient flux, and aquatic ecosystem metabolism. Water Resources Research, 53, 4606–4623.
Wiley Online LibraryCASWeb of Science®Google Scholar - Weigelhofer, G., Hein, T., & Bondar-Kunze, E. (2018). Phosphorus and nitrogen dynamics in riverine systems: Human impacts and management options. In S. Schmutz & J. Sendzimir (Eds.), Riverine ecosystem management: Aquatic ecology series 8. Cham, Switzerland: Springer. https://doi.org/10.1007/978-3-319-73250-3_10
CrossrefGoogle Scholar - Weirich, J. J. (2021). Beaver moderated fire resistance in the north cascades and potential for climate change adaptation (Masters thesis collection 660). Eastern Washington University. https://dc.ewu.edu/theses/660
Google Scholar - Westbrook, C. J. (2021). Beaver as agents of plant disturbance. In Plant disturbance ecology (pp. 489–528). Elsevier.
CrossrefGoogle Scholar - Westbrook, C. J., Ronnquist, A., & Bedard-Haughn, A. (2020). Hydrological functioning of a beaver dam sequence and regional dam persistence during an extreme rainstorm. Hydrological Processes, 34(18), 3726–3737.
Wiley Online LibraryWeb of Science®Google Scholar - Westbrook, C. J., Cooper, D. J., & Baker, B. W. (2006). Beaver dams and overbank floods influence groundwater–surface water interactions of a Rocky Mountain Riparian area. Water Resources Research, 42(6), 06404. https://doi.org/10.1029/2005WR004560
Wiley Online LibraryWeb of Science®Google Scholar - Wheaton, J. M., Bennett, S. N., Bouwes, N. W., Maestas, J. D., & Shahverdian, S. M. (2019). Low-tech process-based restoration of Riverscapes: Design manual. Utah State University Restoration Consortium.
Google Scholar - Whipple, A. (2019). Riparian resilience in the face of interacting disturbances: Understanding complex interactions between wildfire, erosion, and beaver (Castor canadensis) in grazed dryland riparian systems of low order streams in North Central Washington State, USA (Masters thesis collection 586). Eastern Washington University.
Google Scholar - Williams, A. P., Abatzoglou, J. T., Gershunov, A., Guzman-Morales, J., Bishop, D. A., Balch, J. K., & Lettenmaier, D. P. (2019). Observed impacts of anthropogenic climate change on wildfire in California. Earth’s Future, 7(8), 892–910.
Wiley Online LibraryWeb of Science®Google Scholar - Williams, J. (2013). Exploring the onset of high-impact mega-fires through a forest land management prism. Forest Ecology and Management, 294, 4–10.
CrossrefWeb of Science®Google Scholar - Wohl, E. (2013). Landscape-scale carbon storage associated with beaver dams. Geophysical Research Letters, 40(14), 3631–3636.
Wiley Online LibraryCASWeb of Science®Google Scholar - Wohl, E. (2021a). An integrative conceptualization of floodplain storage. Reviews of Geophysics, 59(2), e2020RG000724.
Wiley Online LibraryWeb of Science®Google Scholar - Wohl, E. (2021b). Legacy effects of loss of beavers in the continental United States. Environmental Research Letters, 16(2), 025010.
CrossrefWeb of Science®Google Scholar - Wohl, E., Castro, J., Cluer, B., Merritts, D., Powers, P., Staab, B., & Thorne, C. (2021). Rediscovering, reevaluating, and restoring lost river-wetland corridors. Frontiers in Earth Science, 9, 511.
CrossrefWeb of Science®Google Scholar - Wohl, E., Lininger, K. B., & Scott, D. N. (2017). River beads as a conceptual framework for building carbon storage and resilience to extreme climate events into river management. Biogeochemistry, 141(3), 365–383.
CrossrefWeb of Science®Google Scholar - Wohl, E., Marshall, A. E., Scamardo, J., White, D., & Morrison, R. R. (2022). Biogeomorphic influences on river corridor resilience to wildfire disturbances in a mountain stream of the Southern Rockies, USA. Science of the Total Environment, 820, 153321.
CrossrefCASPubMedWeb of Science®Google Scholar - Yousaf, A., Khalid, N., Aqeel, M., Noman, A., Naeem, N., Sarfraz, W., Ejaz, U., Qaiser, Z., & Khalid, A. (2021). Nitrogen dynamics in wetland systems and its impact on biodiversity. Nitrogen, 2, 196–217. https://doi.org/10.3390/nitrogen2020013
CrossrefCASGoogle Scholar